
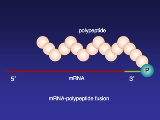
MRNA display
Encyclopedia
mRNA display is a display technique used for in vitro protein
, and/or peptide
evolution to create molecules that can bind to a desired target. The process results in translated peptides or proteins that are associated with their mRNA progenitor via a puromycin
linkage. The complex then binds to an immobilized target in a selection step (affinity chromatography
). The mRNA-protein fusions that bind well are then reverse transcribed to cDNA and their sequence amplified via a polymerase chain reaction
. The end result is a nucleotide
sequence that encodes a peptide with high affinity for the molecule of interest.
Puromycin
is an analogue of the 3’ end of a tyrosyl-tRNA with a part of its structure mimics a molecule of adenosine, and the other part mimics a molecule of tyrosine
. Compared to the cleavable ester bond in a tyrosyl-tRNA, puromycin has a non-hydrolysable amide bond. As a result, puromycin interferes with translation, and causes premature release of translation products.
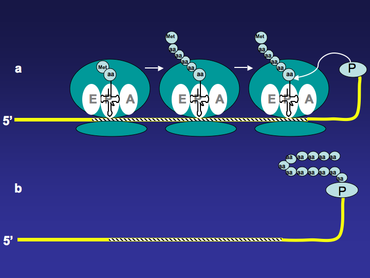 All mRNA templates used for mRNA display technology have puromycin at their 3’ end. As translation proceeds, ribosome moves along the mRNA template, and once it reaches the 3’ end of the template, the fused puromycin will enter ribosome’s A site and be incorporated into the nascent peptide. The mRNA-polypeptide fusion is then released from the ribosome (Figure 2).
All mRNA templates used for mRNA display technology have puromycin at their 3’ end. As translation proceeds, ribosome moves along the mRNA template, and once it reaches the 3’ end of the template, the fused puromycin will enter ribosome’s A site and be incorporated into the nascent peptide. The mRNA-polypeptide fusion is then released from the ribosome (Figure 2).
To synthesize an mRNA-polypeptide fusion, the fused puromycin is not the only modification to the mRNA template. Oligonucletides and other spacers need to be recruited along with the puromycin to provide flexibility and proper length for the puromycin to enter the A site. Ideally, the linker between the 3’ end of an mRNA and the puromycin has to be flexible and long enough to allow the puromycin to enter the A site upon translation of the last codon. This enables the efficient production of high-quality, full-length mRNA-polypeptide fusion. Rihe Liu et al. optimized the 3’-puromycin oligonucleotide spacer. They reported that dA25 in combination with a Spacer 9 (Glen Research), and dAdCdCP at the 5’ terminus worked the best for the fusion reaction. They found that linkers longer than 40 nucleotides and shorter than 16 nucleotides showed greatly reduced efficiency of fusion formation. Also, when the sequence rUrUP presented adjacent to the puromycin, fusion did not form efficiently .
In addition to providing flexibility and length, the poly dA portion of the linker also allows further purification of the mRNA-polypeptide fusion due to its high affinity for dT cellulose resin.
The mRNA-polypeptide fusions can be selected over immobilized selection targets for several rounds with increasing stringency. After each round of selection, those library members that stay bound to the immobilized target are PCR amplified, and non-binders are washed off.
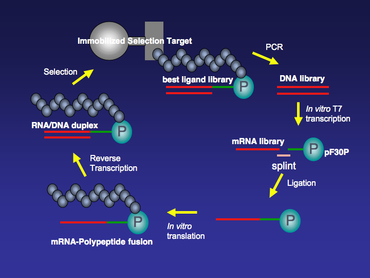
 The synthesis of an mRNA display library starts from the synthesis of a DNA library. A DNA library for any protein or small peptide of interest can be synthesized by solid-phase synthesis followed by PCR amplification. Usually, each member of this DNA library has a T7 RNA polymerase transcription site and a ribosomal binding site at the 5’ end. The T7 promoter region allows large-scale in vitro T7 transcription to transcribe the DNA library into an mRNA library, which provides templates for the in vitro translation reaction later. The ribosomal binding site in the 5’-untranslated region (5’ UTR) is designed according to the in vitro translation system to be used. There are two popular commercially available in vitro translation systems. One is E. Coli S30 Extract System (Promega) that requires a Shine-Dalgarno sequence in the 5’ UTR as a ribosomal binding site; the other one is Red Nova Lysate (Novagen), which needs a ΔTMV ribosomal binding site.
The synthesis of an mRNA display library starts from the synthesis of a DNA library. A DNA library for any protein or small peptide of interest can be synthesized by solid-phase synthesis followed by PCR amplification. Usually, each member of this DNA library has a T7 RNA polymerase transcription site and a ribosomal binding site at the 5’ end. The T7 promoter region allows large-scale in vitro T7 transcription to transcribe the DNA library into an mRNA library, which provides templates for the in vitro translation reaction later. The ribosomal binding site in the 5’-untranslated region (5’ UTR) is designed according to the in vitro translation system to be used. There are two popular commercially available in vitro translation systems. One is E. Coli S30 Extract System (Promega) that requires a Shine-Dalgarno sequence in the 5’ UTR as a ribosomal binding site; the other one is Red Nova Lysate (Novagen), which needs a ΔTMV ribosomal binding site.
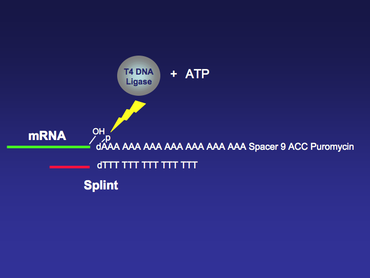
Once the mRNA library is generated, it will be Urea-PAGE purified and ligated using T4 DNA ligase
to the DNA spacer linker containing puromycin at the 3’ end. In this ligation step, a piece of mRNA is ligated with a single stranded DNA with the help from T4 DNA ligase. This is not a standard T4 DNA ligase ligation reaction, where two pieces of double stranded DNA are ligated together. To increase the yield of this special ligation, a single stranded DNA splint may be used to aid the ligation reaction. The 5’ terminus of the splint is designed to be complementary to the 3’ end of the mRNA, and the 3’ terminus of the splint is designed to be complementary to the 5’ end of the DNA spacer linker, which usually consists of poly dA nucleotides (Figure 3).
 The ligated mRNA-DNA-puromycin library is translated in Red Nova Lysate (Novagen) or E. Coli S30 Extract System (Promega), resulting in polypeptides covalently linked in cis to the encoding mRNA. The in vitro translation can also be done in a PURE (protein synthesis using recombinant elements) system. PURE system is an E. Coli cell-free translation system in which only essential translation components are present. Some components, such as amino acids and aminoacyl-tRNA synthases (AARSs) can be omitted from the system. Instead, chemically acylated tRNA can be added into the PURE system. It has been shown that some unnatural amino acids, such as N-methyl-amino acid accylated tRNA can be incorporated into peptides or mRNA-polypeptide fusions in a PURE system .
The ligated mRNA-DNA-puromycin library is translated in Red Nova Lysate (Novagen) or E. Coli S30 Extract System (Promega), resulting in polypeptides covalently linked in cis to the encoding mRNA. The in vitro translation can also be done in a PURE (protein synthesis using recombinant elements) system. PURE system is an E. Coli cell-free translation system in which only essential translation components are present. Some components, such as amino acids and aminoacyl-tRNA synthases (AARSs) can be omitted from the system. Instead, chemically acylated tRNA can be added into the PURE system. It has been shown that some unnatural amino acids, such as N-methyl-amino acid accylated tRNA can be incorporated into peptides or mRNA-polypeptide fusions in a PURE system .
After translation, the single-stranded mRNA portions of the fusions will be converted to heteroduplex of RNA/DNA by reverse transcriptase
to eliminate any unwanted RNA secondary structures, and render the nucleic acid portion of the fusion more stable. This step is a standard reverse transcription reaction. For instance, it can be done by using Superscript II (GIBCO-BRL) following the manufacturer’s protocol.
The mRNA/DNA-polypeptide fusions can be selected over immobilized selection targets for several rounds (Figure 4). There might be a relatively high background for the first few rounds of selection, and this can be minimized by increasing selection stringency, such as adjusting salt concentration, amount of detergent, and/or temperature during the target/fusion binding period. Following binding selection, those library members that stay bound to the immobilized target are PCR amplified. The PCR amplification step will enrich the population from the mRNA-display library that has higher affinity for the immobilized target. Error-prone PCR can also be done in between each round of selection to further increase the diversity of the mRNA-display library and reduce background in selection .
, bacterial display
, yeast display
, and ribosome display
, mRNA display technology has many advantages over the others . The first three biological display libraries listed have polypeptides or proteins expressed on the respective microorganism’s cell surface and the accompanying coding information for each polypeptide or protein is retrievable from the microorganism’s genome. However, the library size for these three in vivo display systems is limited by the transformation efficiency of each organism. For example, the library size for phage and bacterial display is limited to 1-10 × 10^9 different members. The library size for yeast display is even smaller. Moreover, these cell-based display system only allow the screening and enrichment of peptides/proteins containing natural amino acids. In contrast, mRNA display and ribosomse display are in vitro selection methods. They allow a library size as large as 10^15 different members. The large library size increases the probability to select very rare sequences, and also improves the diversity of the selected sequences. In addition, in vitro selection methods remove unwanted selection pressure, such as poor protein expression, and rapid protein degradation, which may reduce the diversity of the selected sequences. Finally, in vitro selection methods allow the application of in vitro mutagenesis and recombination techniques throughout the selection process.
Although both ribosome display and mRNA display are in vitro selection methods, mRNA display has some advantage over the ribosome display technology . mRNA display utilizes covalent mRNA-polypeptide complexes linked through puromycin; whereas, ribosome display utilizes stalled, noncovalent ribosome-mRNA-polypeptide complexes. For ribosome display, selection stringency is limited to keep ribosome-mRNA-polypeptide in a complex because of the noncovalent ribosome-mRNA-polypeptide complexes. This may cause difficulties in reducing background binding during the selection cycle. Also, the polypeptides under selection in a ribosome display system are attached to an enormous rRNA-protein complex, a ribosome, which has a molecular weight of more than 2,000,000 Da. There might be some unpredictable interaction between the selection target and the ribosome, and this may lead to a loss of potential binders during the selection cycle. In contrast, the puromycin DNA spacer linker used in mRNA display technology is much smaller comparing to a ribosome. This linker may have less chance to interact with an immobilized selection target. Thus, mRNA display technology is more likely to give less biased results.
Nine years later, Fukuda and colleagues chose mRNA display method for in vitro evolution of single-chain Fv (scFv) antibody fragments . They selected six different scFv mutants with five consensus mutations. However, kinetic analysis of these mutants showed that their antigen-specificity remained similar to that of the wild type. However, they have demonstrated that two of the five consensus mutations were within the complementarity determining regions (CDRs). And they concluded that mRNA display has the potential for rapid artificial evolution of high-affinity diagnostic and therapeutic antibodies by optimizing their CDRs.
Roberts and coworkers have demonstrated that unnatural peptide oligomers consisting of an N-substituted amino acid can be synthesized as mRNA-polypeptide fusions . N-substituted amino acid-containing peptides have been associated with good proteolytic stability and improved pharmacokinetic properties. This work indicates that mRNA display technology has the potential for selecting drug-like peptides for therapeutic usage resistant to proteolysis.
, and/or peptide
evolution to create molecules that can bind to a desired target. The process results in translated peptides or proteins that are associated with their mRNA progenitor via a puromycin
linkage. The complex then binds to an immobilized target in a selection step (affinity chromatography
). The mRNA-protein fusions that bind well are then reverse transcribed to cDNA and their sequence amplified via a polymerase chain reaction
. The end result is a nucleotide
sequence that encodes a peptide with high affinity for the molecule of interest.
Puromycin
is an analogue of the 3’ end of a tyrosyl-tRNA with a part of its structure mimics a molecule of adenosine, and the other part mimics a molecule of tyrosine
. Compared to the cleavable ester bond in a tyrosyl-tRNA, puromycin has a non-hydrolysable amide bond. As a result, puromycin interferes with translation, and causes premature release of translation products.
All mRNA templates used for mRNA display technology have puromycin at their 3’ end. As translation proceeds, ribosome moves along the mRNA template, and once it reaches the 3’ end of the template, the fused puromycin will enter ribosome’s A site and be incorporated into the nascent peptide. The mRNA-polypeptide fusion is then released from the ribosome (Figure 2).
To synthesize an mRNA-polypeptide fusion, the fused puromycin is not the only modification to the mRNA template. Oligonucletides and other spacers need to be recruited along with the puromycin to provide flexibility and proper length for the puromycin to enter the A site. Ideally, the linker between the 3’ end of an mRNA and the puromycin has to be flexible and long enough to allow the puromycin to enter the A site upon translation of the last codon. This enables the efficient production of high-quality, full-length mRNA-polypeptide fusion. Rihe Liu et al. optimized the 3’-puromycin oligonucleotide spacer. They reported that dA25 in combination with a Spacer 9 (Glen Research), and dAdCdCP at the 5’ terminus worked the best for the fusion reaction. They found that linkers longer than 40 nucleotides and shorter than 16 nucleotides showed greatly reduced efficiency of fusion formation. Also, when the sequence rUrUP presented adjacent to the puromycin, fusion did not form efficiently .
In addition to providing flexibility and length, the poly dA portion of the linker also allows further purification of the mRNA-polypeptide fusion due to its high affinity for dT cellulose resin.
The mRNA-polypeptide fusions can be selected over immobilized selection targets for several rounds with increasing stringency. After each round of selection, those library members that stay bound to the immobilized target are PCR amplified, and non-binders are washed off.
The synthesis of an mRNA display library starts from the synthesis of a DNA library. A DNA library for any protein or small peptide of interest can be synthesized by solid-phase synthesis followed by PCR amplification. Usually, each member of this DNA library has a T7 RNA polymerase transcription site and a ribosomal binding site at the 5’ end. The T7 promoter region allows large-scale in vitro T7 transcription to transcribe the DNA library into an mRNA library, which provides templates for the in vitro translation reaction later. The ribosomal binding site in the 5’-untranslated region (5’ UTR) is designed according to the in vitro translation system to be used. There are two popular commercially available in vitro translation systems. One is E. Coli S30 Extract System (Promega) that requires a Shine-Dalgarno sequence in the 5’ UTR as a ribosomal binding site; the other one is Red Nova Lysate (Novagen), which needs a ΔTMV ribosomal binding site.
Once the mRNA library is generated, it will be Urea-PAGE purified and ligated using T4 DNA ligase
to the DNA spacer linker containing puromycin at the 3’ end. In this ligation step, a piece of mRNA is ligated with a single stranded DNA with the help from T4 DNA ligase. This is not a standard T4 DNA ligase ligation reaction, where two pieces of double stranded DNA are ligated together. To increase the yield of this special ligation, a single stranded DNA splint may be used to aid the ligation reaction. The 5’ terminus of the splint is designed to be complementary to the 3’ end of the mRNA, and the 3’ terminus of the splint is designed to be complementary to the 5’ end of the DNA spacer linker, which usually consists of poly dA nucleotides (Figure 3).
The ligated mRNA-DNA-puromycin library is translated in Red Nova Lysate (Novagen) or E. Coli S30 Extract System (Promega), resulting in polypeptides covalently linked in cis to the encoding mRNA. The in vitro translation can also be done in a PURE (protein synthesis using recombinant elements) system. PURE system is an E. Coli cell-free translation system in which only essential translation components are present. Some components, such as amino acids and aminoacyl-tRNA synthases (AARSs) can be omitted from the system. Instead, chemically acylated tRNA can be added into the PURE system. It has been shown that some unnatural amino acids, such as N-methyl-amino acid accylated tRNA can be incorporated into peptides or mRNA-polypeptide fusions in a PURE system .
After translation, the single-stranded mRNA portions of the fusions will be converted to heteroduplex of RNA/DNA by reverse transcriptase
to eliminate any unwanted RNA secondary structures, and render the nucleic acid portion of the fusion more stable. This step is a standard reverse transcription reaction. For instance, it can be done by using Superscript II (GIBCO-BRL) following the manufacturer’s protocol.
The mRNA/DNA-polypeptide fusions can be selected over immobilized selection targets for several rounds (Figure 4). There might be a relatively high background for the first few rounds of selection, and this can be minimized by increasing selection stringency, such as adjusting salt concentration, amount of detergent, and/or temperature during the target/fusion binding period. Following binding selection, those library members that stay bound to the immobilized target are PCR amplified. The PCR amplification step will enrich the population from the mRNA-display library that has higher affinity for the immobilized target. Error-prone PCR can also be done in between each round of selection to further increase the diversity of the mRNA-display library and reduce background in selection .
, bacterial display
, yeast display
, and ribosome display
, mRNA display technology has many advantages over the others . The first three biological display libraries listed have polypeptides or proteins expressed on the respective microorganism’s cell surface and the accompanying coding information for each polypeptide or protein is retrievable from the microorganism’s genome. However, the library size for these three in vivo display systems is limited by the transformation efficiency of each organism. For example, the library size for phage and bacterial display is limited to 1-10 × 10^9 different members. The library size for yeast display is even smaller. Moreover, these cell-based display system only allow the screening and enrichment of peptides/proteins containing natural amino acids. In contrast, mRNA display and ribosomse display are in vitro selection methods. They allow a library size as large as 10^15 different members. The large library size increases the probability to select very rare sequences, and also improves the diversity of the selected sequences. In addition, in vitro selection methods remove unwanted selection pressure, such as poor protein expression, and rapid protein degradation, which may reduce the diversity of the selected sequences. Finally, in vitro selection methods allow the application of in vitro mutagenesis and recombination techniques throughout the selection process.
Although both ribosome display and mRNA display are in vitro selection methods, mRNA display has some advantage over the ribosome display technology . mRNA display utilizes covalent mRNA-polypeptide complexes linked through puromycin; whereas, ribosome display utilizes stalled, noncovalent ribosome-mRNA-polypeptide complexes. For ribosome display, selection stringency is limited to keep ribosome-mRNA-polypeptide in a complex because of the noncovalent ribosome-mRNA-polypeptide complexes. This may cause difficulties in reducing background binding during the selection cycle. Also, the polypeptides under selection in a ribosome display system are attached to an enormous rRNA-protein complex, a ribosome, which has a molecular weight of more than 2,000,000 Da. There might be some unpredictable interaction between the selection target and the ribosome, and this may lead to a loss of potential binders during the selection cycle. In contrast, the puromycin DNA spacer linker used in mRNA display technology is much smaller comparing to a ribosome. This linker may have less chance to interact with an immobilized selection target. Thus, mRNA display technology is more likely to give less biased results.
Nine years later, Fukuda and colleagues chose mRNA display method for in vitro evolution of single-chain Fv (scFv) antibody fragments . They selected six different scFv mutants with five consensus mutations. However, kinetic analysis of these mutants showed that their antigen-specificity remained similar to that of the wild type. However, they have demonstrated that two of the five consensus mutations were within the complementarity determining regions (CDRs). And they concluded that mRNA display has the potential for rapid artificial evolution of high-affinity diagnostic and therapeutic antibodies by optimizing their CDRs.
Roberts and coworkers have demonstrated that unnatural peptide oligomers consisting of an N-substituted amino acid can be synthesized as mRNA-polypeptide fusions . N-substituted amino acid-containing peptides have been associated with good proteolytic stability and improved pharmacokinetic properties. This work indicates that mRNA display technology has the potential for selecting drug-like peptides for therapeutic usage resistant to proteolysis.
, and/or peptide
evolution to create molecules that can bind to a desired target. The process results in translated peptides or proteins that are associated with their mRNA progenitor via a puromycin
linkage. The complex then binds to an immobilized target in a selection step (affinity chromatography
). The mRNA-protein fusions that bind well are then reverse transcribed to cDNA and their sequence amplified via a polymerase chain reaction
. The end result is a nucleotide
sequence that encodes a peptide with high affinity for the molecule of interest.
Puromycin
is an analogue of the 3’ end of a tyrosyl-tRNA with a part of its structure mimics a molecule of adenosine, and the other part mimics a molecule of tyrosine
. Compared to the cleavable ester bond in a tyrosyl-tRNA, puromycin has a non-hydrolysable amide bond. As a result, puromycin interferes with translation, and causes premature release of translation products.
All mRNA templates used for mRNA display technology have puromycin at their 3’ end. As translation proceeds, ribosome moves along the mRNA template, and once it reaches the 3’ end of the template, the fused puromycin will enter ribosome’s A site and be incorporated into the nascent peptide. The mRNA-polypeptide fusion is then released from the ribosome (Figure 2).
To synthesize an mRNA-polypeptide fusion, the fused puromycin is not the only modification to the mRNA template. Oligonucletides and other spacers need to be recruited along with the puromycin to provide flexibility and proper length for the puromycin to enter the A site. Ideally, the linker between the 3’ end of an mRNA and the puromycin has to be flexible and long enough to allow the puromycin to enter the A site upon translation of the last codon. This enables the efficient production of high-quality, full-length mRNA-polypeptide fusion. Rihe Liu et al. optimized the 3’-puromycin oligonucleotide spacer. They reported that dA25 in combination with a Spacer 9 (Glen Research), and dAdCdCP at the 5’ terminus worked the best for the fusion reaction. They found that linkers longer than 40 nucleotides and shorter than 16 nucleotides showed greatly reduced efficiency of fusion formation. Also, when the sequence rUrUP presented adjacent to the puromycin, fusion did not form efficiently .
In addition to providing flexibility and length, the poly dA portion of the linker also allows further purification of the mRNA-polypeptide fusion due to its high affinity for dT cellulose resin.
The mRNA-polypeptide fusions can be selected over immobilized selection targets for several rounds with increasing stringency. After each round of selection, those library members that stay bound to the immobilized target are PCR amplified, and non-binders are washed off.
The synthesis of an mRNA display library starts from the synthesis of a DNA library. A DNA library for any protein or small peptide of interest can be synthesized by solid-phase synthesis followed by PCR amplification. Usually, each member of this DNA library has a T7 RNA polymerase transcription site and a ribosomal binding site at the 5’ end. The T7 promoter region allows large-scale in vitro T7 transcription to transcribe the DNA library into an mRNA library, which provides templates for the in vitro translation reaction later. The ribosomal binding site in the 5’-untranslated region (5’ UTR) is designed according to the in vitro translation system to be used. There are two popular commercially available in vitro translation systems. One is E. Coli S30 Extract System (Promega) that requires a Shine-Dalgarno sequence in the 5’ UTR as a ribosomal binding site; the other one is Red Nova Lysate (Novagen), which needs a ΔTMV ribosomal binding site.
Once the mRNA library is generated, it will be Urea-PAGE purified and ligated using T4 DNA ligase
to the DNA spacer linker containing puromycin at the 3’ end. In this ligation step, a piece of mRNA is ligated with a single stranded DNA with the help from T4 DNA ligase. This is not a standard T4 DNA ligase ligation reaction, where two pieces of double stranded DNA are ligated together. To increase the yield of this special ligation, a single stranded DNA splint may be used to aid the ligation reaction. The 5’ terminus of the splint is designed to be complementary to the 3’ end of the mRNA, and the 3’ terminus of the splint is designed to be complementary to the 5’ end of the DNA spacer linker, which usually consists of poly dA nucleotides (Figure 3).
The ligated mRNA-DNA-puromycin library is translated in Red Nova Lysate (Novagen) or E. Coli S30 Extract System (Promega), resulting in polypeptides covalently linked in cis to the encoding mRNA. The in vitro translation can also be done in a PURE (protein synthesis using recombinant elements) system. PURE system is an E. Coli cell-free translation system in which only essential translation components are present. Some components, such as amino acids and aminoacyl-tRNA synthases (AARSs) can be omitted from the system. Instead, chemically acylated tRNA can be added into the PURE system. It has been shown that some unnatural amino acids, such as N-methyl-amino acid accylated tRNA can be incorporated into peptides or mRNA-polypeptide fusions in a PURE system .
After translation, the single-stranded mRNA portions of the fusions will be converted to heteroduplex of RNA/DNA by reverse transcriptase
to eliminate any unwanted RNA secondary structures, and render the nucleic acid portion of the fusion more stable. This step is a standard reverse transcription reaction. For instance, it can be done by using Superscript II (GIBCO-BRL) following the manufacturer’s protocol.
The mRNA/DNA-polypeptide fusions can be selected over immobilized selection targets for several rounds (Figure 4). There might be a relatively high background for the first few rounds of selection, and this can be minimized by increasing selection stringency, such as adjusting salt concentration, amount of detergent, and/or temperature during the target/fusion binding period. Following binding selection, those library members that stay bound to the immobilized target are PCR amplified. The PCR amplification step will enrich the population from the mRNA-display library that has higher affinity for the immobilized target. Error-prone PCR can also be done in between each round of selection to further increase the diversity of the mRNA-display library and reduce background in selection .
, bacterial display
, yeast display
, and ribosome display
, mRNA display technology has many advantages over the others . The first three biological display libraries listed have polypeptides or proteins expressed on the respective microorganism’s cell surface and the accompanying coding information for each polypeptide or protein is retrievable from the microorganism’s genome. However, the library size for these three in vivo display systems is limited by the transformation efficiency of each organism. For example, the library size for phage and bacterial display is limited to 1-10 × 10^9 different members. The library size for yeast display is even smaller. Moreover, these cell-based display system only allow the screening and enrichment of peptides/proteins containing natural amino acids. In contrast, mRNA display and ribosomse display are in vitro selection methods. They allow a library size as large as 10^15 different members. The large library size increases the probability to select very rare sequences, and also improves the diversity of the selected sequences. In addition, in vitro selection methods remove unwanted selection pressure, such as poor protein expression, and rapid protein degradation, which may reduce the diversity of the selected sequences. Finally, in vitro selection methods allow the application of in vitro mutagenesis and recombination techniques throughout the selection process.
Although both ribosome display and mRNA display are in vitro selection methods, mRNA display has some advantage over the ribosome display technology . mRNA display utilizes covalent mRNA-polypeptide complexes linked through puromycin; whereas, ribosome display utilizes stalled, noncovalent ribosome-mRNA-polypeptide complexes. For ribosome display, selection stringency is limited to keep ribosome-mRNA-polypeptide in a complex because of the noncovalent ribosome-mRNA-polypeptide complexes. This may cause difficulties in reducing background binding during the selection cycle. Also, the polypeptides under selection in a ribosome display system are attached to an enormous rRNA-protein complex, a ribosome, which has a molecular weight of more than 2,000,000 Da. There might be some unpredictable interaction between the selection target and the ribosome, and this may lead to a loss of potential binders during the selection cycle. In contrast, the puromycin DNA spacer linker used in mRNA display technology is much smaller comparing to a ribosome. This linker may have less chance to interact with an immobilized selection target. Thus, mRNA display technology is more likely to give less biased results.
Nine years later, Fukuda and colleagues chose mRNA display method for in vitro evolution of single-chain Fv (scFv) antibody fragments . They selected six different scFv mutants with five consensus mutations. However, kinetic analysis of these mutants showed that their antigen-specificity remained similar to that of the wild type. However, they have demonstrated that two of the five consensus mutations were within the complementarity determining regions (CDRs). And they concluded that mRNA display has the potential for rapid artificial evolution of high-affinity diagnostic and therapeutic antibodies by optimizing their CDRs.
Roberts and coworkers have demonstrated that unnatural peptide oligomers consisting of an N-substituted amino acid can be synthesized as mRNA-polypeptide fusions . N-substituted amino acid-containing peptides have been associated with good proteolytic stability and improved pharmacokinetic properties. This work indicates that mRNA display technology has the potential for selecting drug-like peptides for therapeutic usage resistant to proteolysis.
Protein
Proteins are biochemical compounds consisting of one or more polypeptides typically folded into a globular or fibrous form, facilitating a biological function. A polypeptide is a single linear polymer chain of amino acids bonded together by peptide bonds between the carboxyl and amino groups of...
, and/or peptide
Peptide
Peptides are short polymers of amino acid monomers linked by peptide bonds. They are distinguished from proteins on the basis of size, typically containing less than 50 monomer units. The shortest peptides are dipeptides, consisting of two amino acids joined by a single peptide bond...
evolution to create molecules that can bind to a desired target. The process results in translated peptides or proteins that are associated with their mRNA progenitor via a puromycin
Puromycin
Puromycin is an antibiotic that is a protein synthesis inhibitor by inhibiting translation.-Inhibition of translation:Puromycin is an aminonucleoside antibiotic, derived from the Streptomyces alboniger bacterium, that causes premature chain termination during translation taking place in the...
linkage. The complex then binds to an immobilized target in a selection step (affinity chromatography
Affinity chromatography
Affinity chromatography is a method of separating biochemical mixtures and based on a highly specific interaction such as that between antigen and antibody, enzyme and substrate, or receptor and ligand.-Uses:Affinity chromatography can be used to:...
). The mRNA-protein fusions that bind well are then reverse transcribed to cDNA and their sequence amplified via a polymerase chain reaction
Polymerase chain reaction
The polymerase chain reaction is a scientific technique in molecular biology to amplify a single or a few copies of a piece of DNA across several orders of magnitude, generating thousands to millions of copies of a particular DNA sequence....
. The end result is a nucleotide
Nucleotide
Nucleotides are molecules that, when joined together, make up the structural units of RNA and DNA. In addition, nucleotides participate in cellular signaling , and are incorporated into important cofactors of enzymatic reactions...
sequence that encodes a peptide with high affinity for the molecule of interest.
Puromycin
Puromycin
Puromycin is an antibiotic that is a protein synthesis inhibitor by inhibiting translation.-Inhibition of translation:Puromycin is an aminonucleoside antibiotic, derived from the Streptomyces alboniger bacterium, that causes premature chain termination during translation taking place in the...
is an analogue of the 3’ end of a tyrosyl-tRNA with a part of its structure mimics a molecule of adenosine, and the other part mimics a molecule of tyrosine
Tyrosine
Tyrosine or 4-hydroxyphenylalanine, is one of the 22 amino acids that are used by cells to synthesize proteins. Its codons are UAC and UAU. It is a non-essential amino acid with a polar side group...
. Compared to the cleavable ester bond in a tyrosyl-tRNA, puromycin has a non-hydrolysable amide bond. As a result, puromycin interferes with translation, and causes premature release of translation products.
To synthesize an mRNA-polypeptide fusion, the fused puromycin is not the only modification to the mRNA template. Oligonucletides and other spacers need to be recruited along with the puromycin to provide flexibility and proper length for the puromycin to enter the A site. Ideally, the linker between the 3’ end of an mRNA and the puromycin has to be flexible and long enough to allow the puromycin to enter the A site upon translation of the last codon. This enables the efficient production of high-quality, full-length mRNA-polypeptide fusion. Rihe Liu et al. optimized the 3’-puromycin oligonucleotide spacer. They reported that dA25 in combination with a Spacer 9 (Glen Research), and dAdCdCP at the 5’ terminus worked the best for the fusion reaction. They found that linkers longer than 40 nucleotides and shorter than 16 nucleotides showed greatly reduced efficiency of fusion formation. Also, when the sequence rUrUP presented adjacent to the puromycin, fusion did not form efficiently .
In addition to providing flexibility and length, the poly dA portion of the linker also allows further purification of the mRNA-polypeptide fusion due to its high affinity for dT cellulose resin.
The mRNA-polypeptide fusions can be selected over immobilized selection targets for several rounds with increasing stringency. After each round of selection, those library members that stay bound to the immobilized target are PCR amplified, and non-binders are washed off.
Method
Once the mRNA library is generated, it will be Urea-PAGE purified and ligated using T4 DNA ligase
DNA ligase
In molecular biology, DNA ligase is a specific type of enzyme, a ligase, that repairs single-stranded discontinuities in double stranded DNA molecules, in simple words strands that have double-strand break . Purified DNA ligase is used in gene cloning to join DNA molecules together...
to the DNA spacer linker containing puromycin at the 3’ end. In this ligation step, a piece of mRNA is ligated with a single stranded DNA with the help from T4 DNA ligase. This is not a standard T4 DNA ligase ligation reaction, where two pieces of double stranded DNA are ligated together. To increase the yield of this special ligation, a single stranded DNA splint may be used to aid the ligation reaction. The 5’ terminus of the splint is designed to be complementary to the 3’ end of the mRNA, and the 3’ terminus of the splint is designed to be complementary to the 5’ end of the DNA spacer linker, which usually consists of poly dA nucleotides (Figure 3).
After translation, the single-stranded mRNA portions of the fusions will be converted to heteroduplex of RNA/DNA by reverse transcriptase
Reverse transcriptase
In the fields of molecular biology and biochemistry, a reverse transcriptase, also known as RNA-dependent DNA polymerase, is a DNA polymerase enzyme that transcribes single-stranded RNA into single-stranded DNA. It also helps in the formation of a double helix DNA once the RNA has been reverse...
to eliminate any unwanted RNA secondary structures, and render the nucleic acid portion of the fusion more stable. This step is a standard reverse transcription reaction. For instance, it can be done by using Superscript II (GIBCO-BRL) following the manufacturer’s protocol.
The mRNA/DNA-polypeptide fusions can be selected over immobilized selection targets for several rounds (Figure 4). There might be a relatively high background for the first few rounds of selection, and this can be minimized by increasing selection stringency, such as adjusting salt concentration, amount of detergent, and/or temperature during the target/fusion binding period. Following binding selection, those library members that stay bound to the immobilized target are PCR amplified. The PCR amplification step will enrich the population from the mRNA-display library that has higher affinity for the immobilized target. Error-prone PCR can also be done in between each round of selection to further increase the diversity of the mRNA-display library and reduce background in selection .
Advantage
Although there are many other molecular display technologies, such as phage displayPhage display
Phage display is a method for the study of protein–protein, protein–peptide, and protein–DNA interactions that uses bacteriophages to connect proteins with the genetic information that encodes them. Phage Display was originally invented by George P...
, bacterial display
Bacterial display
Bacterial display is a protein engineering technique used for in vitro protein evolution...
, yeast display
Yeast display
Yeast display is a technique used in the field of protein engineering. The yeast display technique was first published by the laboratory of Professor K. Dane Wittrup. The technology was sold to Abbott Laboratories in 2001....
, and ribosome display
Ribosome display
Ribosome display is a technique used to perform in vitro protein evolution to create proteins that can bind to a desired ligand. The process results in translated proteins that are associated with their mRNA progenitor which is used, as a complex, to bind to an immobilized ligand in a selection step...
, mRNA display technology has many advantages over the others . The first three biological display libraries listed have polypeptides or proteins expressed on the respective microorganism’s cell surface and the accompanying coding information for each polypeptide or protein is retrievable from the microorganism’s genome. However, the library size for these three in vivo display systems is limited by the transformation efficiency of each organism. For example, the library size for phage and bacterial display is limited to 1-10 × 10^9 different members. The library size for yeast display is even smaller. Moreover, these cell-based display system only allow the screening and enrichment of peptides/proteins containing natural amino acids. In contrast, mRNA display and ribosomse display are in vitro selection methods. They allow a library size as large as 10^15 different members. The large library size increases the probability to select very rare sequences, and also improves the diversity of the selected sequences. In addition, in vitro selection methods remove unwanted selection pressure, such as poor protein expression, and rapid protein degradation, which may reduce the diversity of the selected sequences. Finally, in vitro selection methods allow the application of in vitro mutagenesis and recombination techniques throughout the selection process.
Although both ribosome display and mRNA display are in vitro selection methods, mRNA display has some advantage over the ribosome display technology . mRNA display utilizes covalent mRNA-polypeptide complexes linked through puromycin; whereas, ribosome display utilizes stalled, noncovalent ribosome-mRNA-polypeptide complexes. For ribosome display, selection stringency is limited to keep ribosome-mRNA-polypeptide in a complex because of the noncovalent ribosome-mRNA-polypeptide complexes. This may cause difficulties in reducing background binding during the selection cycle. Also, the polypeptides under selection in a ribosome display system are attached to an enormous rRNA-protein complex, a ribosome, which has a molecular weight of more than 2,000,000 Da. There might be some unpredictable interaction between the selection target and the ribosome, and this may lead to a loss of potential binders during the selection cycle. In contrast, the puromycin DNA spacer linker used in mRNA display technology is much smaller comparing to a ribosome. This linker may have less chance to interact with an immobilized selection target. Thus, mRNA display technology is more likely to give less biased results.
Application
In 1997, Roberts and Szostak showed that fusions between a synthetic mRNA and its encoded myc epitope could be enriched from a pool of random sequence mRNA-polypeptide fusions by immunoprecipitation .Nine years later, Fukuda and colleagues chose mRNA display method for in vitro evolution of single-chain Fv (scFv) antibody fragments . They selected six different scFv mutants with five consensus mutations. However, kinetic analysis of these mutants showed that their antigen-specificity remained similar to that of the wild type. However, they have demonstrated that two of the five consensus mutations were within the complementarity determining regions (CDRs). And they concluded that mRNA display has the potential for rapid artificial evolution of high-affinity diagnostic and therapeutic antibodies by optimizing their CDRs.
Roberts and coworkers have demonstrated that unnatural peptide oligomers consisting of an N-substituted amino acid can be synthesized as mRNA-polypeptide fusions . N-substituted amino acid-containing peptides have been associated with good proteolytic stability and improved pharmacokinetic properties. This work indicates that mRNA display technology has the potential for selecting drug-like peptides for therapeutic usage resistant to proteolysis.
External links
mRNA display is a display technique used for in vitro proteinProtein
Proteins are biochemical compounds consisting of one or more polypeptides typically folded into a globular or fibrous form, facilitating a biological function. A polypeptide is a single linear polymer chain of amino acids bonded together by peptide bonds between the carboxyl and amino groups of...
, and/or peptide
Peptide
Peptides are short polymers of amino acid monomers linked by peptide bonds. They are distinguished from proteins on the basis of size, typically containing less than 50 monomer units. The shortest peptides are dipeptides, consisting of two amino acids joined by a single peptide bond...
evolution to create molecules that can bind to a desired target. The process results in translated peptides or proteins that are associated with their mRNA progenitor via a puromycin
Puromycin
Puromycin is an antibiotic that is a protein synthesis inhibitor by inhibiting translation.-Inhibition of translation:Puromycin is an aminonucleoside antibiotic, derived from the Streptomyces alboniger bacterium, that causes premature chain termination during translation taking place in the...
linkage. The complex then binds to an immobilized target in a selection step (affinity chromatography
Affinity chromatography
Affinity chromatography is a method of separating biochemical mixtures and based on a highly specific interaction such as that between antigen and antibody, enzyme and substrate, or receptor and ligand.-Uses:Affinity chromatography can be used to:...
). The mRNA-protein fusions that bind well are then reverse transcribed to cDNA and their sequence amplified via a polymerase chain reaction
Polymerase chain reaction
The polymerase chain reaction is a scientific technique in molecular biology to amplify a single or a few copies of a piece of DNA across several orders of magnitude, generating thousands to millions of copies of a particular DNA sequence....
. The end result is a nucleotide
Nucleotide
Nucleotides are molecules that, when joined together, make up the structural units of RNA and DNA. In addition, nucleotides participate in cellular signaling , and are incorporated into important cofactors of enzymatic reactions...
sequence that encodes a peptide with high affinity for the molecule of interest.
Puromycin
Puromycin
Puromycin is an antibiotic that is a protein synthesis inhibitor by inhibiting translation.-Inhibition of translation:Puromycin is an aminonucleoside antibiotic, derived from the Streptomyces alboniger bacterium, that causes premature chain termination during translation taking place in the...
is an analogue of the 3’ end of a tyrosyl-tRNA with a part of its structure mimics a molecule of adenosine, and the other part mimics a molecule of tyrosine
Tyrosine
Tyrosine or 4-hydroxyphenylalanine, is one of the 22 amino acids that are used by cells to synthesize proteins. Its codons are UAC and UAU. It is a non-essential amino acid with a polar side group...
. Compared to the cleavable ester bond in a tyrosyl-tRNA, puromycin has a non-hydrolysable amide bond. As a result, puromycin interferes with translation, and causes premature release of translation products.
To synthesize an mRNA-polypeptide fusion, the fused puromycin is not the only modification to the mRNA template. Oligonucletides and other spacers need to be recruited along with the puromycin to provide flexibility and proper length for the puromycin to enter the A site. Ideally, the linker between the 3’ end of an mRNA and the puromycin has to be flexible and long enough to allow the puromycin to enter the A site upon translation of the last codon. This enables the efficient production of high-quality, full-length mRNA-polypeptide fusion. Rihe Liu et al. optimized the 3’-puromycin oligonucleotide spacer. They reported that dA25 in combination with a Spacer 9 (Glen Research), and dAdCdCP at the 5’ terminus worked the best for the fusion reaction. They found that linkers longer than 40 nucleotides and shorter than 16 nucleotides showed greatly reduced efficiency of fusion formation. Also, when the sequence rUrUP presented adjacent to the puromycin, fusion did not form efficiently .
In addition to providing flexibility and length, the poly dA portion of the linker also allows further purification of the mRNA-polypeptide fusion due to its high affinity for dT cellulose resin.
The mRNA-polypeptide fusions can be selected over immobilized selection targets for several rounds with increasing stringency. After each round of selection, those library members that stay bound to the immobilized target are PCR amplified, and non-binders are washed off.
Method
Once the mRNA library is generated, it will be Urea-PAGE purified and ligated using T4 DNA ligase
DNA ligase
In molecular biology, DNA ligase is a specific type of enzyme, a ligase, that repairs single-stranded discontinuities in double stranded DNA molecules, in simple words strands that have double-strand break . Purified DNA ligase is used in gene cloning to join DNA molecules together...
to the DNA spacer linker containing puromycin at the 3’ end. In this ligation step, a piece of mRNA is ligated with a single stranded DNA with the help from T4 DNA ligase. This is not a standard T4 DNA ligase ligation reaction, where two pieces of double stranded DNA are ligated together. To increase the yield of this special ligation, a single stranded DNA splint may be used to aid the ligation reaction. The 5’ terminus of the splint is designed to be complementary to the 3’ end of the mRNA, and the 3’ terminus of the splint is designed to be complementary to the 5’ end of the DNA spacer linker, which usually consists of poly dA nucleotides (Figure 3).
After translation, the single-stranded mRNA portions of the fusions will be converted to heteroduplex of RNA/DNA by reverse transcriptase
Reverse transcriptase
In the fields of molecular biology and biochemistry, a reverse transcriptase, also known as RNA-dependent DNA polymerase, is a DNA polymerase enzyme that transcribes single-stranded RNA into single-stranded DNA. It also helps in the formation of a double helix DNA once the RNA has been reverse...
to eliminate any unwanted RNA secondary structures, and render the nucleic acid portion of the fusion more stable. This step is a standard reverse transcription reaction. For instance, it can be done by using Superscript II (GIBCO-BRL) following the manufacturer’s protocol.
The mRNA/DNA-polypeptide fusions can be selected over immobilized selection targets for several rounds (Figure 4). There might be a relatively high background for the first few rounds of selection, and this can be minimized by increasing selection stringency, such as adjusting salt concentration, amount of detergent, and/or temperature during the target/fusion binding period. Following binding selection, those library members that stay bound to the immobilized target are PCR amplified. The PCR amplification step will enrich the population from the mRNA-display library that has higher affinity for the immobilized target. Error-prone PCR can also be done in between each round of selection to further increase the diversity of the mRNA-display library and reduce background in selection .
Advantage
Although there are many other molecular display technologies, such as phage displayPhage display
Phage display is a method for the study of protein–protein, protein–peptide, and protein–DNA interactions that uses bacteriophages to connect proteins with the genetic information that encodes them. Phage Display was originally invented by George P...
, bacterial display
Bacterial display
Bacterial display is a protein engineering technique used for in vitro protein evolution...
, yeast display
Yeast display
Yeast display is a technique used in the field of protein engineering. The yeast display technique was first published by the laboratory of Professor K. Dane Wittrup. The technology was sold to Abbott Laboratories in 2001....
, and ribosome display
Ribosome display
Ribosome display is a technique used to perform in vitro protein evolution to create proteins that can bind to a desired ligand. The process results in translated proteins that are associated with their mRNA progenitor which is used, as a complex, to bind to an immobilized ligand in a selection step...
, mRNA display technology has many advantages over the others . The first three biological display libraries listed have polypeptides or proteins expressed on the respective microorganism’s cell surface and the accompanying coding information for each polypeptide or protein is retrievable from the microorganism’s genome. However, the library size for these three in vivo display systems is limited by the transformation efficiency of each organism. For example, the library size for phage and bacterial display is limited to 1-10 × 10^9 different members. The library size for yeast display is even smaller. Moreover, these cell-based display system only allow the screening and enrichment of peptides/proteins containing natural amino acids. In contrast, mRNA display and ribosomse display are in vitro selection methods. They allow a library size as large as 10^15 different members. The large library size increases the probability to select very rare sequences, and also improves the diversity of the selected sequences. In addition, in vitro selection methods remove unwanted selection pressure, such as poor protein expression, and rapid protein degradation, which may reduce the diversity of the selected sequences. Finally, in vitro selection methods allow the application of in vitro mutagenesis and recombination techniques throughout the selection process.
Although both ribosome display and mRNA display are in vitro selection methods, mRNA display has some advantage over the ribosome display technology . mRNA display utilizes covalent mRNA-polypeptide complexes linked through puromycin; whereas, ribosome display utilizes stalled, noncovalent ribosome-mRNA-polypeptide complexes. For ribosome display, selection stringency is limited to keep ribosome-mRNA-polypeptide in a complex because of the noncovalent ribosome-mRNA-polypeptide complexes. This may cause difficulties in reducing background binding during the selection cycle. Also, the polypeptides under selection in a ribosome display system are attached to an enormous rRNA-protein complex, a ribosome, which has a molecular weight of more than 2,000,000 Da. There might be some unpredictable interaction between the selection target and the ribosome, and this may lead to a loss of potential binders during the selection cycle. In contrast, the puromycin DNA spacer linker used in mRNA display technology is much smaller comparing to a ribosome. This linker may have less chance to interact with an immobilized selection target. Thus, mRNA display technology is more likely to give less biased results.
Application
In 1997, Roberts and Szostak showed that fusions between a synthetic mRNA and its encoded myc epitope could be enriched from a pool of random sequence mRNA-polypeptide fusions by immunoprecipitation .Nine years later, Fukuda and colleagues chose mRNA display method for in vitro evolution of single-chain Fv (scFv) antibody fragments . They selected six different scFv mutants with five consensus mutations. However, kinetic analysis of these mutants showed that their antigen-specificity remained similar to that of the wild type. However, they have demonstrated that two of the five consensus mutations were within the complementarity determining regions (CDRs). And they concluded that mRNA display has the potential for rapid artificial evolution of high-affinity diagnostic and therapeutic antibodies by optimizing their CDRs.
Roberts and coworkers have demonstrated that unnatural peptide oligomers consisting of an N-substituted amino acid can be synthesized as mRNA-polypeptide fusions . N-substituted amino acid-containing peptides have been associated with good proteolytic stability and improved pharmacokinetic properties. This work indicates that mRNA display technology has the potential for selecting drug-like peptides for therapeutic usage resistant to proteolysis.
External links
mRNA display is a display technique used for in vitro proteinProtein
Proteins are biochemical compounds consisting of one or more polypeptides typically folded into a globular or fibrous form, facilitating a biological function. A polypeptide is a single linear polymer chain of amino acids bonded together by peptide bonds between the carboxyl and amino groups of...
, and/or peptide
Peptide
Peptides are short polymers of amino acid monomers linked by peptide bonds. They are distinguished from proteins on the basis of size, typically containing less than 50 monomer units. The shortest peptides are dipeptides, consisting of two amino acids joined by a single peptide bond...
evolution to create molecules that can bind to a desired target. The process results in translated peptides or proteins that are associated with their mRNA progenitor via a puromycin
Puromycin
Puromycin is an antibiotic that is a protein synthesis inhibitor by inhibiting translation.-Inhibition of translation:Puromycin is an aminonucleoside antibiotic, derived from the Streptomyces alboniger bacterium, that causes premature chain termination during translation taking place in the...
linkage. The complex then binds to an immobilized target in a selection step (affinity chromatography
Affinity chromatography
Affinity chromatography is a method of separating biochemical mixtures and based on a highly specific interaction such as that between antigen and antibody, enzyme and substrate, or receptor and ligand.-Uses:Affinity chromatography can be used to:...
). The mRNA-protein fusions that bind well are then reverse transcribed to cDNA and their sequence amplified via a polymerase chain reaction
Polymerase chain reaction
The polymerase chain reaction is a scientific technique in molecular biology to amplify a single or a few copies of a piece of DNA across several orders of magnitude, generating thousands to millions of copies of a particular DNA sequence....
. The end result is a nucleotide
Nucleotide
Nucleotides are molecules that, when joined together, make up the structural units of RNA and DNA. In addition, nucleotides participate in cellular signaling , and are incorporated into important cofactors of enzymatic reactions...
sequence that encodes a peptide with high affinity for the molecule of interest.
Puromycin
Puromycin
Puromycin is an antibiotic that is a protein synthesis inhibitor by inhibiting translation.-Inhibition of translation:Puromycin is an aminonucleoside antibiotic, derived from the Streptomyces alboniger bacterium, that causes premature chain termination during translation taking place in the...
is an analogue of the 3’ end of a tyrosyl-tRNA with a part of its structure mimics a molecule of adenosine, and the other part mimics a molecule of tyrosine
Tyrosine
Tyrosine or 4-hydroxyphenylalanine, is one of the 22 amino acids that are used by cells to synthesize proteins. Its codons are UAC and UAU. It is a non-essential amino acid with a polar side group...
. Compared to the cleavable ester bond in a tyrosyl-tRNA, puromycin has a non-hydrolysable amide bond. As a result, puromycin interferes with translation, and causes premature release of translation products.
To synthesize an mRNA-polypeptide fusion, the fused puromycin is not the only modification to the mRNA template. Oligonucletides and other spacers need to be recruited along with the puromycin to provide flexibility and proper length for the puromycin to enter the A site. Ideally, the linker between the 3’ end of an mRNA and the puromycin has to be flexible and long enough to allow the puromycin to enter the A site upon translation of the last codon. This enables the efficient production of high-quality, full-length mRNA-polypeptide fusion. Rihe Liu et al. optimized the 3’-puromycin oligonucleotide spacer. They reported that dA25 in combination with a Spacer 9 (Glen Research), and dAdCdCP at the 5’ terminus worked the best for the fusion reaction. They found that linkers longer than 40 nucleotides and shorter than 16 nucleotides showed greatly reduced efficiency of fusion formation. Also, when the sequence rUrUP presented adjacent to the puromycin, fusion did not form efficiently .
In addition to providing flexibility and length, the poly dA portion of the linker also allows further purification of the mRNA-polypeptide fusion due to its high affinity for dT cellulose resin.
The mRNA-polypeptide fusions can be selected over immobilized selection targets for several rounds with increasing stringency. After each round of selection, those library members that stay bound to the immobilized target are PCR amplified, and non-binders are washed off.
Method
Once the mRNA library is generated, it will be Urea-PAGE purified and ligated using T4 DNA ligase
DNA ligase
In molecular biology, DNA ligase is a specific type of enzyme, a ligase, that repairs single-stranded discontinuities in double stranded DNA molecules, in simple words strands that have double-strand break . Purified DNA ligase is used in gene cloning to join DNA molecules together...
to the DNA spacer linker containing puromycin at the 3’ end. In this ligation step, a piece of mRNA is ligated with a single stranded DNA with the help from T4 DNA ligase. This is not a standard T4 DNA ligase ligation reaction, where two pieces of double stranded DNA are ligated together. To increase the yield of this special ligation, a single stranded DNA splint may be used to aid the ligation reaction. The 5’ terminus of the splint is designed to be complementary to the 3’ end of the mRNA, and the 3’ terminus of the splint is designed to be complementary to the 5’ end of the DNA spacer linker, which usually consists of poly dA nucleotides (Figure 3).
After translation, the single-stranded mRNA portions of the fusions will be converted to heteroduplex of RNA/DNA by reverse transcriptase
Reverse transcriptase
In the fields of molecular biology and biochemistry, a reverse transcriptase, also known as RNA-dependent DNA polymerase, is a DNA polymerase enzyme that transcribes single-stranded RNA into single-stranded DNA. It also helps in the formation of a double helix DNA once the RNA has been reverse...
to eliminate any unwanted RNA secondary structures, and render the nucleic acid portion of the fusion more stable. This step is a standard reverse transcription reaction. For instance, it can be done by using Superscript II (GIBCO-BRL) following the manufacturer’s protocol.
The mRNA/DNA-polypeptide fusions can be selected over immobilized selection targets for several rounds (Figure 4). There might be a relatively high background for the first few rounds of selection, and this can be minimized by increasing selection stringency, such as adjusting salt concentration, amount of detergent, and/or temperature during the target/fusion binding period. Following binding selection, those library members that stay bound to the immobilized target are PCR amplified. The PCR amplification step will enrich the population from the mRNA-display library that has higher affinity for the immobilized target. Error-prone PCR can also be done in between each round of selection to further increase the diversity of the mRNA-display library and reduce background in selection .
Advantage
Although there are many other molecular display technologies, such as phage displayPhage display
Phage display is a method for the study of protein–protein, protein–peptide, and protein–DNA interactions that uses bacteriophages to connect proteins with the genetic information that encodes them. Phage Display was originally invented by George P...
, bacterial display
Bacterial display
Bacterial display is a protein engineering technique used for in vitro protein evolution...
, yeast display
Yeast display
Yeast display is a technique used in the field of protein engineering. The yeast display technique was first published by the laboratory of Professor K. Dane Wittrup. The technology was sold to Abbott Laboratories in 2001....
, and ribosome display
Ribosome display
Ribosome display is a technique used to perform in vitro protein evolution to create proteins that can bind to a desired ligand. The process results in translated proteins that are associated with their mRNA progenitor which is used, as a complex, to bind to an immobilized ligand in a selection step...
, mRNA display technology has many advantages over the others . The first three biological display libraries listed have polypeptides or proteins expressed on the respective microorganism’s cell surface and the accompanying coding information for each polypeptide or protein is retrievable from the microorganism’s genome. However, the library size for these three in vivo display systems is limited by the transformation efficiency of each organism. For example, the library size for phage and bacterial display is limited to 1-10 × 10^9 different members. The library size for yeast display is even smaller. Moreover, these cell-based display system only allow the screening and enrichment of peptides/proteins containing natural amino acids. In contrast, mRNA display and ribosomse display are in vitro selection methods. They allow a library size as large as 10^15 different members. The large library size increases the probability to select very rare sequences, and also improves the diversity of the selected sequences. In addition, in vitro selection methods remove unwanted selection pressure, such as poor protein expression, and rapid protein degradation, which may reduce the diversity of the selected sequences. Finally, in vitro selection methods allow the application of in vitro mutagenesis and recombination techniques throughout the selection process.
Although both ribosome display and mRNA display are in vitro selection methods, mRNA display has some advantage over the ribosome display technology . mRNA display utilizes covalent mRNA-polypeptide complexes linked through puromycin; whereas, ribosome display utilizes stalled, noncovalent ribosome-mRNA-polypeptide complexes. For ribosome display, selection stringency is limited to keep ribosome-mRNA-polypeptide in a complex because of the noncovalent ribosome-mRNA-polypeptide complexes. This may cause difficulties in reducing background binding during the selection cycle. Also, the polypeptides under selection in a ribosome display system are attached to an enormous rRNA-protein complex, a ribosome, which has a molecular weight of more than 2,000,000 Da. There might be some unpredictable interaction between the selection target and the ribosome, and this may lead to a loss of potential binders during the selection cycle. In contrast, the puromycin DNA spacer linker used in mRNA display technology is much smaller comparing to a ribosome. This linker may have less chance to interact with an immobilized selection target. Thus, mRNA display technology is more likely to give less biased results.
Application
In 1997, Roberts and Szostak showed that fusions between a synthetic mRNA and its encoded myc epitope could be enriched from a pool of random sequence mRNA-polypeptide fusions by immunoprecipitation .Nine years later, Fukuda and colleagues chose mRNA display method for in vitro evolution of single-chain Fv (scFv) antibody fragments . They selected six different scFv mutants with five consensus mutations. However, kinetic analysis of these mutants showed that their antigen-specificity remained similar to that of the wild type. However, they have demonstrated that two of the five consensus mutations were within the complementarity determining regions (CDRs). And they concluded that mRNA display has the potential for rapid artificial evolution of high-affinity diagnostic and therapeutic antibodies by optimizing their CDRs.
Roberts and coworkers have demonstrated that unnatural peptide oligomers consisting of an N-substituted amino acid can be synthesized as mRNA-polypeptide fusions . N-substituted amino acid-containing peptides have been associated with good proteolytic stability and improved pharmacokinetic properties. This work indicates that mRNA display technology has the potential for selecting drug-like peptides for therapeutic usage resistant to proteolysis.