

Species-area curve
Encyclopedia
In ecology
, a species-area curve is a relationship between the area of a habitat, or of part of a habitat, and the number of species found within that area. Larger areas tend to contain larger numbers of species, and empirically, the relative numbers seem to follow systematic mathematical relationships. The species-area relationship is usually constructed for a single type of organism, such as all vascular plants or all species of a specific trophic level
within a particular site. It is rarely, if ever, constructed for all types of organisms if simply because of the prodigious data requirements. It is related to, but not identical with, the species discovery curve
.
Ecologists have proposed a wide range of factors determining the slope and elevation of the species-area relationship. These factors include the relative balance between immigration and extinction, rate and magnitude of disturbance on small vs. large areas, predator-prey dynamics, and clustering of individuals of the same species as a result of dispersal limitation or habitat heterogeneity. The species-area relationship has been reputed to follow from the 2nd law of thermodynamics
. In contrast to these "mechanistic" explanations, others assert the need to test whether the pattern is simply the result of a random sampling process.
Authors have classified the species-area relationship according to the type of habitats being sampled and the census design used. Frank Preston, an early investigator of the theory of the species-area relationship, divided it into two types: samples (a census of a contiguous habitat that grows in census area, also called "mainland" species-area relationships), and isolates (a census of discontiguous habitats, such as islands, also called "island" species-area relationships). Michael Rosenzweig also notes that species-area relationships for very large areas—those collecting different biogeographic provinces or continents—behave differently from species-area relationships from islands or smaller contiguous areas. It has been presumed that "island"-like species-area relationships have higher slopes (in log-log space) than "mainland" relationships, but a recent metaanalysis of almost 700 species-area relationships found the former had lower slopes than the latter.
Regardless of census design and habitat type, species-area relationships are often fit with a simple function. Frank Preston advocated the power function based on his investigation of the lognormal species-abundance distribution. If S is the number of species, A is area, c is a constant (number of species in the smallest sampling area) and z is the slope of the species area relationship in log-log space, then the power function species-area relationship goes as:

which looks like a straight line on log-log axes
. In contrast, Henry Gleason
championed the semilog model:
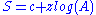
which looks like a straight line on semilog axes
, where area is logged and the number of species is arithmetic. In either case, the species-area relationship is almost always decelerating (has a negative second derivative) when plotted arithmetically.
Species-area relationships are often graphed for islands (or habitats that are otherwise isolated from one another, such as woodlots in an agricultural landscape) of different sizes. Although larger islands tend to have more species, it is possible that a smaller island will have more than a larger one. In contrast, species-area relationships for contiguous habitats will always rise as areas increases, provided that the sample plots are nested within one another.
The species-area relationship for mainland areas (contiguous habitats) will differ according to the census design used to construct it. A common method is to use quadrats of successively larger size, so that the area enclosed by each one includes the area enclosed by the smaller one (i.e. areas are nested).
In the first part of the 20th century plant ecologists often used the species-area curve to estimate the minimum size of a quadrat necessary to adequately characterize a community. This is done by plotting the curve (usually on arithmetic axes, not log-log or semilog axes), and estimating the area after which using larger quadrats results in the addition of only a few more species. This is called the minimal area. A quadrat that encloses the minimal area is called a relevé, and using species-area curves in this way is called the relevé method. It was largely developed by the Swiss
ecologist Josias Braun-Blanquet
.
Estimation of the minimal area from the curve is necessarily subjective, so some authors prefer to define minimal area as the area enclosing at least 95 percent (or some other large proportion) of the total species found. The problem with this is that the species area curve does not usually approach an asymptote
, so it is not obvious what should be taken as the total. In fact, the number of species always increases with area up to the point where the area of the entire world has been accumulated.
Ecology
Ecology is the scientific study of the relations that living organisms have with respect to each other and their natural environment. Variables of interest to ecologists include the composition, distribution, amount , number, and changing states of organisms within and among ecosystems...
, a species-area curve is a relationship between the area of a habitat, or of part of a habitat, and the number of species found within that area. Larger areas tend to contain larger numbers of species, and empirically, the relative numbers seem to follow systematic mathematical relationships. The species-area relationship is usually constructed for a single type of organism, such as all vascular plants or all species of a specific trophic level
Trophic level
The trophic level of an organism is the position it occupies in a food chain. The word trophic derives from the Greek τροφή referring to food or feeding. A food chain represents a succession of organisms that eat another organism and are, in turn, eaten themselves. The number of steps an organism...
within a particular site. It is rarely, if ever, constructed for all types of organisms if simply because of the prodigious data requirements. It is related to, but not identical with, the species discovery curve
Species discovery curve
In ecology, the species discovery curve is a graph recording the cumulative number of species of living things recorded in a particular environment as a function of the cumulative effort expended searching for them...
.
Ecologists have proposed a wide range of factors determining the slope and elevation of the species-area relationship. These factors include the relative balance between immigration and extinction, rate and magnitude of disturbance on small vs. large areas, predator-prey dynamics, and clustering of individuals of the same species as a result of dispersal limitation or habitat heterogeneity. The species-area relationship has been reputed to follow from the 2nd law of thermodynamics
Second law of thermodynamics
The second law of thermodynamics is an expression of the tendency that over time, differences in temperature, pressure, and chemical potential equilibrate in an isolated physical system. From the state of thermodynamic equilibrium, the law deduced the principle of the increase of entropy and...
. In contrast to these "mechanistic" explanations, others assert the need to test whether the pattern is simply the result of a random sampling process.
Authors have classified the species-area relationship according to the type of habitats being sampled and the census design used. Frank Preston, an early investigator of the theory of the species-area relationship, divided it into two types: samples (a census of a contiguous habitat that grows in census area, also called "mainland" species-area relationships), and isolates (a census of discontiguous habitats, such as islands, also called "island" species-area relationships). Michael Rosenzweig also notes that species-area relationships for very large areas—those collecting different biogeographic provinces or continents—behave differently from species-area relationships from islands or smaller contiguous areas. It has been presumed that "island"-like species-area relationships have higher slopes (in log-log space) than "mainland" relationships, but a recent metaanalysis of almost 700 species-area relationships found the former had lower slopes than the latter.
Regardless of census design and habitat type, species-area relationships are often fit with a simple function. Frank Preston advocated the power function based on his investigation of the lognormal species-abundance distribution. If S is the number of species, A is area, c is a constant (number of species in the smallest sampling area) and z is the slope of the species area relationship in log-log space, then the power function species-area relationship goes as:
which looks like a straight line on log-log axes
Log-log graph
In science and engineering, a log-log graph or log-log plot is a two-dimensional graph of numerical data that uses logarithmic scales on both the horizontal and vertical axes...
. In contrast, Henry Gleason
Henry Gleason
Henry Allan Gleason was a noted American ecologist, botanist, and taxonomist, most recognized for his endorsement of the individualistic/open community concept of ecological succession.- Life and work :...
championed the semilog model:
which looks like a straight line on semilog axes
Semilog graph
In science and engineering, a semi-log graph or semi-log plot is a way of visualizing data that are changing with an exponential relationship. One axis is plotted on a logarithmic scale...
, where area is logged and the number of species is arithmetic. In either case, the species-area relationship is almost always decelerating (has a negative second derivative) when plotted arithmetically.
Species-area relationships are often graphed for islands (or habitats that are otherwise isolated from one another, such as woodlots in an agricultural landscape) of different sizes. Although larger islands tend to have more species, it is possible that a smaller island will have more than a larger one. In contrast, species-area relationships for contiguous habitats will always rise as areas increases, provided that the sample plots are nested within one another.
The species-area relationship for mainland areas (contiguous habitats) will differ according to the census design used to construct it. A common method is to use quadrats of successively larger size, so that the area enclosed by each one includes the area enclosed by the smaller one (i.e. areas are nested).
In the first part of the 20th century plant ecologists often used the species-area curve to estimate the minimum size of a quadrat necessary to adequately characterize a community. This is done by plotting the curve (usually on arithmetic axes, not log-log or semilog axes), and estimating the area after which using larger quadrats results in the addition of only a few more species. This is called the minimal area. A quadrat that encloses the minimal area is called a relevé, and using species-area curves in this way is called the relevé method. It was largely developed by the Swiss
Switzerland
Switzerland name of one of the Swiss cantons. ; ; ; or ), in its full name the Swiss Confederation , is a federal republic consisting of 26 cantons, with Bern as the seat of the federal authorities. The country is situated in Western Europe,Or Central Europe depending on the definition....
ecologist Josias Braun-Blanquet
Josias Braun-Blanquet
Josias Braun-Blanquet was an influential phytosociologist and botanist. Braun-Blanquet was born in Chur, Switzerland and died in Montpellier, France.-Phytosociology:...
.
Estimation of the minimal area from the curve is necessarily subjective, so some authors prefer to define minimal area as the area enclosing at least 95 percent (or some other large proportion) of the total species found. The problem with this is that the species area curve does not usually approach an asymptote
Asymptote
In analytic geometry, an asymptote of a curve is a line such that the distance between the curve and the line approaches zero as they tend to infinity. Some sources include the requirement that the curve may not cross the line infinitely often, but this is unusual for modern authors...
, so it is not obvious what should be taken as the total. In fact, the number of species always increases with area up to the point where the area of the entire world has been accumulated.
See also
- Scaling pattern of occupancyScaling pattern of occupancyIn spatial ecology and macroecology, scaling pattern of occupancy , also known as the area-of-occupancy is the way in which species distribution changes across spatial scales. In physical geography and image analysis, it is similar to the modifiable areal unit problem. Simon A...
- Species richnessSpecies richnessSpecies richness is the number of different species in a given area. It is represented in equation form as S.Species richness is the fundamental unit in which to assess the homogeneity of an environment. Typically, species richness is used in conservation studies to determine the sensitivity of...
- Storage effectStorage effectThe storage effect is a coexistence mechanism proposed in the ecological theory of species coexistence, which tries to explain how such a wide variety of similar species are able to coexist within the same ecological community or guild...
- Unified neutral theory of biodiversityUnified neutral theory of biodiversityThe unified neutral theory of biodiversity and biogeography is a hypothesis and the title of a monograph by ecologist Stephen Hubbell...
(UNTB)